
Click on image to see full-size!
- the battery uses a chemical reaction to produce electrical power
- The motor converts electricity into rotary movement.
- The transmission adapts the rotary movement to the appropriate speed and power
- The trigger controls the electricity to the motor
- The chuck holds the bit in place.
- The bit drills the whole
Click on image to see full-size!
- Domains can be 25 to 500 residues long; most are less than 200 residues.
- The average protein contains 2 or 3 domains.
- The total number of different types of domains ~1000 - 3000
- Each domain has a characteristic sequence, which folds into a characteristic secondary and tertiary structure.
- Each domain therefore has a characteristic shape which enables it to perform a function.
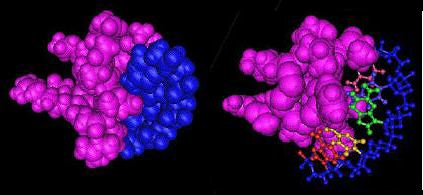
- Fig. 1. A space-filling model (ZnF domain in pink, DNA in blue) showing complementary shapes and binding of the protein domain and ligand.
- Fig. 2. DNA rendered as a ball-and-stick model, with sugar-phosphate backbone in blue, and nitrogenous bases distinguished by color.
- Cadherin domains. Cadherin domains are extracellular regions which mediate binding between cell adhesion proteins on the surface of adjacent cells.
- Basic Leucine zipper domain (bZIP domain). The bZIP domain mediates sequence specific binding to DNA.
- IGc Immunoglobulin domain. These domains are involved with dimerization between immunoglobulin chains to produce functional antibodies. They are components of immunoglobulins, T-cell receptors, CD1 cell surface glycoproteins, secretory glycoproteins A/C, and Major Histocompatibility Complex (MHC) class I/II molecules.
- Phosphotyrosine-binding domain (PTB). PTB domains usually bind to phosphorylated tyrosine residues. They are often found in signal transduction proteins.
- Src homology 2 domain (SH2). SH2 domains are often found
in signal transduction proteins. SH2 domains confer binding to phosphorylated
tyrosine (pTyr).

Click on image to see full-size!
- electric drill: battery + motor + transmission + trigger switch + drill bit
- electric saw: battery + motor + transmission + trigger switch + saw blade
- electric sander: battery + motor + transmission + trigger switch + sanding wheel
Individual protein domains can also be used in different combinations.
Each domain retains its distinct structure and function (as does a batter
or motor), but when combined with different domains produces proteins with
different functions. For example, the graphic to the left, shows that the
the Zinc-finger domain (ZnF) occurs with other domains in different combinations
to produce many other proteins with different functions.
An example of the modular function
of protein domains.
Click on image to see full-size!
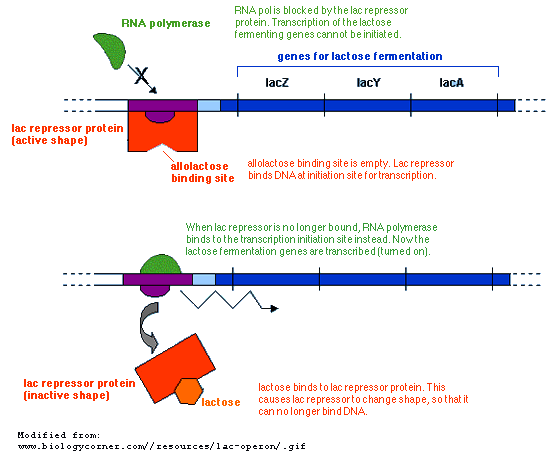
These genes are turned off by a lactose repressor protein, which binds to the DNA upstream of the lac genes, and thereby prevents RNA polymerase from initiating transcription.
However the lac repressor also has a second binding site, which binds allolactose, an isomer of lactose. When lactose is present, the allolactose isomer binds to the lac repressor. This causes the protein to change shape in such a way that the DNA binding site releases from the transcription initiation site.
With the lac repressor gone, RNA polymerase
binds to the initiation site, and begins transcribing the lac genes.
![]()
Click on image to see full-size!
- with the allolactose binding site empty, the DNA-binding site binds to the initiation site for transcription of the lac genes.
- with allolactose bound, the lac repressor releases from the DNA

Click on image to see full-size!
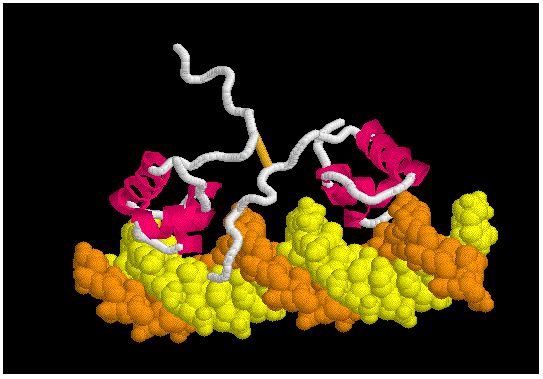
- The lac repressor is a dimer, composed of two identical amino acid chains (left image).
- Each monomer is composed of 3 structural domains as shown in the image to the left.
- The DNA-binding domain (light green or pink) bind to the transcription initiation site in DNA via a helix-turn-helix (HTH) motif.
- Two structural domains (brown and grey) form the Peripla_BP_1 domain. This domain binds sugars. It is found in a family of membrane proteins which function as receptors and transporters. In the lac repressor family, the sugar binding domain binds a sugar which changes the DNA binding activity of the repressor domain .
Click on image to see full-size!
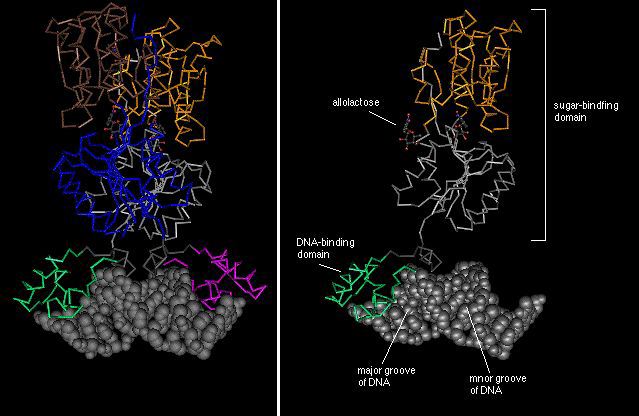
Each monomer is composed of 3 structural domains as shown by the different colors (right image).
The DNA-binding domain (green) binds in the major groove of the DNA molecule.
Two structural domains (brown and grey) form the functional sugar-binding
domain. In the lac repressor, this is the binding site for
allolactose. Two allolactose molecules are depicted as ball-and-stick models.